1. ОСОБЕННОСТИ УСТРОЙСТВА И ФУНКЦИОНИРОВАНИЯ ОБОНЯТЕЛЬНОГО АППАРАТА СОБАКИ
1. ОСОБЕННОСТИ УСТРОЙСТВА И ФУНКЦИОНИРОВАНИЯ ОБОНЯТЕЛЬНОГО АППАРАТА СОБАКИ
Обоняние, т. е. способность воспринимать запахи окружающей среды, играет особую роль в жизни и при дрессировке служебных собак. Собаки относятся к макросматикам, т. е. животным с сильно развитым обонятельным аппаратом, и если у них искусственно отключить зрение и слух, они легко ориентируются в окружающей среде.
Жизнь диких предков собак напрямую зависела от того, насколько успешно они могли ориентироваться по запахам, а также находить по запаховому следу убегающую от них добычу. «Если бы собака писала книгу об органах чувств, то несомненно самая длинная глава была бы посвящена обонянию», — писал Г. Деккер, автор книги, посвященной способностям животных воспринимать окружающий мир.
Данная глава пособия посвящена обонятельной ориентации собак, на базе которой при дрессировке вырабатываются многочисленные навыки, используемые в дальнейшем при работе с собаками.
Периферическая часть обонятельного анализатора собаки своим строением и функцией предназначена для забора запаховой информации, ее первичной обработки, формирования и передачи в центральную нервную систему соответствующего импульса.
Молекулы запахового вещества вместе со струёй воздуха при вдохе попадают в периферический отдел обонятельного анализатора, где химическая энергия запаховых веществ трансформируется в специфический нервный процесс — возбуждение обонятельных рецепторов.
Первоначально воздух проходит через ноздри собаки, имеющие круглую часть и боковые вырезы, затем попадает в носовую полость, где имеются три носовых хода, разделенных складками слизистой оболочки, и три носовые раковины — нижние, средние и верхние. Верхний, средний и нижний носовые ходы соединяются общим носовым ходом. Нижний носовой ход — дыхательный, средний и общий — смешанные, верхний — только обонятельный, он заканчивается слепо. У собак обонятельная область носа резко увеличивается за счет развития системы этмоидальных раковин, представляющих собой выросты решетчатой кости.
При обычном вдохе большая часть вдыхаемого воздуха проходит через нижний, меньшая — через средний и общий носовые ходы и лишь незначительная часть попадает на обонятельный эпителий, выстилающий верхний носовой ход и этмоидальные раковины.
К аппарату обоняния относят также якобсонов орган, расположенный на носовой перегородке недалеко от входа в носовую полость. Считают, что с его помощью собака реагирует на сильные резкие раздражающие запахи, что позволяет ей избежать их попадания на чувствительные участки обонятельного эпителия верхней носовой раковины.
Изучение самого обонятельного эпителия показало, что он содержит рецепторные клетки, которые относятся к наиболее древним — первичночувствующим рецепторам. Они снабжены собственным центральным отростком, выполняющим роль аксона и передающим полученную клеткой информацию в обонятельные луковицы, которые являются первичными центрами обонятельного анализатора.
Количество обонятельных рецепторов весьма велико. В значительной мере оно определяется площадью, занимаемой обонятельным эпителием, хотя зависит также от размеров рецепторов и плотности их расположения в обонятельном эпителии. У немецкой овчарки насчитывается 224 млн. обонятельных клеток при площади обонятельного эпителия 196,46 см2, для сравнения: у человека 10 млн. клеток при площади эпителия 10 см2. Обонятельные клетки имеют веретеновидную форму и снабжены, кроме центрального, коротким периферическим отростком, заканчивающимся на поверхности утолщением — обонятельной булавой с волосками различной длины. Количество волосков в обонятельных клетках собаки обычно более 20, длина большинства их составляет о кол о 15–30 мкм. Отдельные обонятельные клетки снабжены более длинными волосками. В волосках, по всей видимости, осуществляются первичные процессы обонятельной рецепции, и возможно, что волоски обеспечивают увеличение рецептивной поверхности обонятельной клетки.
Рецепторные клетки окружены и изолированы друг от друга телами опорных клеток, часть из которых расположена вблизи базальной мембраны рецепторного слоя. Такие клетки носят название базальных. В соединительнотканном слое обонятельной выстилки локализуются концевые отделы боуменовых желез, главным образом продуцирующих слизь, в которую погружены обонятельные волоски. Здесь же располагаются многочисленные кровеносные сосуды и пучки обонятельных нервов, образованные центральными отростками обонятельных клеток, которые в виде многочисленных коротких и тонких обонятельных нитей проходят через отверстие решетчатой кости к обонятельным луковицам мозга.
При биохимическом исследовании в обонятельной выстилке некоторых животных были обнаружены каратиноиды, однако до сих пор не ясно, имеют ли они непосредственное отношение к обонятельной рецепции.
Центральная часть обонятельного анализатора
Обонятельные луковицы лежат на базальной поверхности лобных долей головного мозга. Они построены по типу корковых центров со сложным расположением элементов и могут быть отнесены к корковым структурам. Здесь осуществляется первое синаптическое переключение для сигналов, приходящих от обонятельных рецепторов.
Сравнение обонятельных проводящих путей с другими сенсорными путями показывает ряд характерных особенностей обонятельной системы. Она не претерпела существен ной перестройки в процессе эволюции позвоночных, у обонятельного анализатора не обнаружено специального представительства в новой коре, как у других сенсорных систем. Ранее считалось, что высший центр обонятельного анализатора представлен обширной областью в старой коре, так называемом «обонятельном мозге», однако последние данные показывают, что собственно обонятельная область более ограничена, а роль ряда структур «обонятельного мозга» выходит далеко за рамки собственно обонятельной функции. Эти структуры связаны с интеграцией соматовегетативных реакций, управлением эмоциональным состоянием, мотивацией поведения и т. д. Их можно рассматривать как ассоциативные обонятельные центры. Становится понятным, почему для многих запахов характерна эмоциональная окраска, наблюдаются многообразные физиологические эффекты (дыхательные, сосудистые) при действии пахучих веществ с приятным и неприятным запахами. Приходящие к «обонятельному мозгу» обонятельные сигналы, коррелируя с другими сенсорными сигналами, служат активаторами, включающими необонятельные типы активности животного — пищевую, половую, оборонительную и т. д., возможно, что обонятельные сигналы в большей степени, чем возбуждения сенсорных систем, способствуют созданию и поддержанию различных мотивационных состояний.
Функционирование обонятельных рецепторов и проблема классификации запахов
Процесс взаимодействия молекул пахучего вещества с обонятельными рецепторами изучен очень слабо. Считается установленным, что для их возбуждения требуется непосредственный контакт молекулы пахучего вещества с рецепторными структурами. Обонятельные рецепторы специализировались в процессе эволюции и приобрели особую чувствительность и специфичность к большой группе низкомолекулярных соединений. Остается неизвестным, какими именно физическими или химическими параметрами определяется эффективность вещества как обонятельного стимула. В качестве необходимых условий указывали на летучесть, достаточную способность к адсорбции на обонятельной выстилке, растворимость в липидах и до некоторой степени в воде, молекулярный вес, лежащий в определенных границах, конфигурацию молекулы, обеспечивающую ее сродство к рецепторам и др.
Различия в ответах разных рецепторных клеток на запахи позволяют допустить существование нескольких (возможно, многих) типов «рецептивных мест», которые по-разному распределены на рецепторах.
Попыток объяснения механизма первичного процесса взаимодействия молекул пахучих веществ с рецептором сделано немало. Они тесно связаны с проблемой классификации запахов, но до сих пор ни одно объяснение не является общепризнанным.
Неоднократно пытались выяснить связь запаха с химическим строением вещества. Для некоторых классов запахов характерно присутствие в молекуле определенных функциональных групп, так называемых «одорифоров». Предполагалось, что именно эти группы проявляют химическое сродство к «рецептивному месту» обонятельного эпителия, однако язык химических формул не позволяет точно предсказать запахи веществ. Существуют несомненные корреляции, ограниченные правила, но всеобщей однозначной зависимости между запахом и химическим строением найти не удалось.
Широкую известность получила «стереохимическая теория запаха» Эймура. Согласно ей все вещества с одинаковыми запахами имеют одинаковую форму или схожие по форме части молекул и могут заполнять определенные «гнезда» («лунки») на поверхности обонятельных клеток. Число таких «гнезд» ограничено и соответствует числу первичных запахов, из которых складываются все остальные. Было выделено около семи первичных запахов: камфорный, эфирный, острый, цветочный, мятный, мускусный, гнилостный. «Лунка» эфирного — вытянутая, камфорного — имеет вид эллиптической чаши, мускусного — сходна, но чуть больше по размеру, цветочного — имеет сложную форму и т. д. Экспериментальная проверка не подтвердила данную концепцию.
Делались попытки связать запах с энергетическими переходами в молекулах вещества. Наиболее известна теория Райта. Он обратил внимание на колебательную и вращательную энергию молекул, которую определяют по спектрам поглощения в инфракрасной области, и хотел выявить связь между запахами веществ и их спектрами в далекой инфракрасной области. По этой гипотезе в обонятельном эпителии имеется несколько типов пигментов, молекулы которых находятся в возбужденном состоянии и обуславливают поляризацию мембраны обонятельных клеток. При совпадении частоты колебания молекулы запахового вещества и молекулы соответствующего пигмента последняя переходит в более стабильное состояние, что вызывает деполяризацию мембраны и первичный потенциал, распространяющийся по поверхности рецепторной клетки. Данная гипотеза также имеет ряд затруднений.
Таким образом, проблема связи качества запаха со свойствами молекул, а также механизмы взаимодействия пахучего вещества с рецепторами еще ждут своего рассмотрения.
Регуляция обонятельной чувствительности
Регуляция обонятельной чувствительности на уровне рецепторов в основном осуществляется посредством изменения условий доступа к ним пахучего вещества, т. е. связана с особенностями дыхания собаки. Как уже упоминалось выше, при нормальном дыхании лишь малая часть воздуха с молекулами запахового вещества достигает обонятельного эпителия. Чтобы улучшить проникновение паров пахучего вещества в обонятельные отделы носа, собака должна сделать глубокий вдох, что обычно происходит при обнюхивании либо короткие частые вдохи.
Поступление запаховых частиц в носовую полость зависит также от строения ноздрей собаки. С. А. Корытин при исследовании обонятельной ориентации псовых показал, что они лучше воспринимают запахи, источник которых находится сбоку, хуже — спереди или сзади от животного. Дальнейшее изучение проблемы дало следующий результат. Воздух поступает в нос собаки через ноздри, имеющие круглую часть и боковые вырезы (рис. 9а), причем при обычном вдохе около 60% воздуха поступает через круглую ноздрю и около 40% — через вырезы; при принюхивании же наоборот — уже около 60% воздуха поступает через боковые вырезы. Контуры струй вдыхаемого воздуха напоминают веер или диск, из которого удалены два сектора — один спереди, другой сзади. Перед носом животного располагается область (около 60% воображаемого диска), откуда воздух в ноздри не засасывается. По 50° влево и вправо от нее, в секторах от 30° до 80° влево и вправо от продольной оси головы воздух поступает через круглую часть ноздри, от 80° до 130° — через боковой вырез носа. Таким образом, боковые вырезы служат для успешной обонятельной ориентации в пространстве, позволяя обследовать воздух сбоку и частично сзади животного.
При нормальном дыхании собаки верхняя и нижняя стенки бокового выреза носа параллельны друг другу. При принюхивании его стенки становятся вогнутыми, наружная щель сужается и боковой вырез представляет собой почти замкнутую подлине трубочку с нешироким продольным разрезом. Так как круглая часть ноздри также сужается, то создаются условия для поступления в нос воздуха преимущественно сбоку и сзади от животного: струи вдыхаемого воздуха представляют собой два сектора уже по 120° (от 30° до 150°) влево и вправо от продольной оси головы (рис. 96).
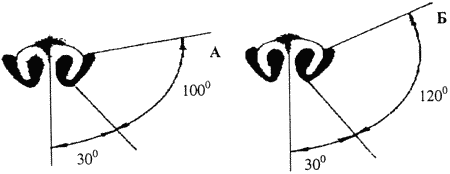
Рис. 9. Контуры струй вдыхаемого собакой воздуха при обычном вдохе (а) и при принюхивании (б)
Плоскость, проведенная через контуры вдыхаемых струй обеих ноздрей, составляет с осью головы угол около 30°, что соответствует направленности боковых вырезов носа (рис. 10 а). Для сравнения: у кошек относительно небольшие вырезы носа направлены вдоль оси головы, а не под углом к ней (рис. 10 б). Известно, что среди кошачьих нет хищников, способных к длительному преследованию жертвы по ее запаховому следу.

Рис. 10. Строение ноздрей собаки (а) и кошки (б)
Наблюдение за собаками, идущими по следу с использованием нижнего чутья, показывает, что плоскость, проведенная через боковые вырезы носа обеих ноздрей, оказывается перпендикулярной земной поверхности. Это позволяет им обследовать горизонтальный столб воздуха, представляющий собой цилиндр диаметром около 15 см, расположенный непосредственно под мордой животного (рис. 11 a). В таком положении собаки делают меньше всего ошибок при проработке следа. Необходимо помнить, что при ротовом дыхании диаметр исследуемого столба воздуха уменьшается за счет изменения струи всасываемого через нос воздуха.
Если собака идет по следу с поднятой головой, ею обследуется горизонтальный столб воздуха меньшего диаметра (рис. 11 б). Также играет роль большее перемешивание воздуха в пределах этого столба, и собака может допустить большее количество ошибок при работе по следу. Принюхиваясь к дальнему источнику запаха, животное поднимает голову, как правило, до уровня, когда плоскость боковых вырезов носа становится почти параллельной поверхности земли (рис. 11 в). Это позволяет ему лучше обследовать горизонтальные потоки воздуха, несущие запаховую информацию от отдельных источников.
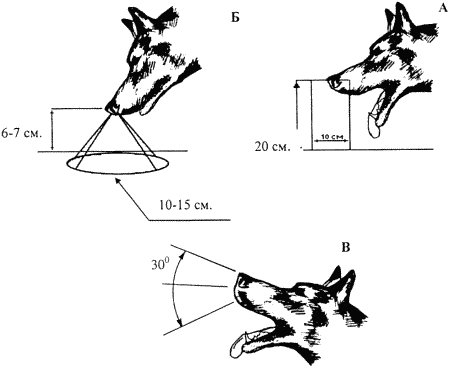
Рис. 11. Структура струй вдыхаемого собакой воздуха при проработке следа (а, б) и при принюхивании к дальнему источнику запаха (в)
Хотелось бы остановиться на вопросе о том, как поступает воздух в носовую полость собаки, когда она движется по следу с достаточной скоростью. Собаки бегут по следу с открытым ртом, дыхание при этом частое и поверхностное: при незначительных нагрузках — около 100–140 вдохов в минуту, при значительных (бег в течение 30 мин. и более) 200–250 вдохов в мин. Охлаждение тела осуществляется за счет дыхания. Через нос проходит лишь 5–15% от вдыхаемого воздуха, причем почти весь он проходит через круглую часть ноздри. При вдохе этот воздух проходит через носовую полость и вместе с основной массой воздуха поступает в легкие (рис. 12 а).
При выдохе через рот также наблюдается засасывание воздуха в ноздри извне (рис. 12 б).
Возможно это связано с тем, что при движении воздуха из легких с большой скоростью в глотке (самом узком месте), создается область пониженного давления, что способствует засасыванию порции воздуха через хоаны из носовой полости. Эта часть воздуха в легкие не попадает, а, пройдя через нос, выбрасывается наружу с основной массой выдыхаемого животными воздуха.
Таким образом, обонятельная информация поступает к бегущей по следу собаке непрерывно — на вдохе и на выдохе, что позволяет ей достаточно эффективно корректировать свое поведение при преследовании убегающего человека или зверя.
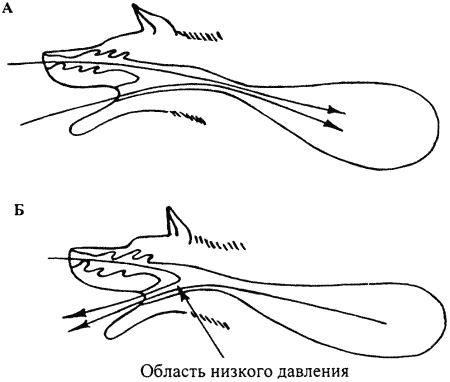
Рис. 12. Прохождение потока воздуха через нос при вдохе (а) и (б) выдохе во время быстрого движения (бега) собаки по запаховому следу
На бегу собака проводит лишь ориентировочный анализ поступающих запахов, возможно этого достаточно для того, чтобы не потерять искомый запах. В случае утери следа собака останавливается и тщательно принюхивается, в основном с закрытым ртом, проводя, таким образом, тщательный анализ поступающих запахов.
Остается открытым вопрос, как собака определяет направление, в котором удалялась добыча. Очевидно, этот сложный нейрофизиологический и поведенческий процесс осуществляется за счет различий во времени при попадании запаховых частиц в левую и правую ноздри, которые увеличиваются из-за веерообразного строения струи вдыхаемого воздуха. Определение направления движения добычи по следу происходит, очевидно, по изменению концентрации запаха на небольшом участке следа.
Возвращаясь к регуляции обонятельной чувствительности, надо отметить, что сами пахучие вещества, попадая в полость носа, вызывают различные дыхательные реакции — учащение, замедление, задержку дыхания. Сужение или расширение сосудов обонятельного эпителия, сдвиги в составе или количестве покрывающей его слизи и другие реакции должны оказывать значительное влияние на функцию обонятельных рецепторов.
Изменение обонятельной функции может быть связано с обонятельной адаптацией, т. е. уменьшением чувствительности к различным запахам после их длительного воздействия, или с сенсибилизацией — повышением чувствительности к какому-либо запаху после соответствующей тренировки или иных воздействий. Эти процессы в определенной степени являются результатом центральной регуляции, иногда выходящими за рамки собственно обонятельной системы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Особенности пищеварения собаки
Особенности пищеварения собаки Собаки являются плотоядными животными, значит, их желудочно-кишечный тракт приспособлен, прежде всего, к перевариванию мясной пищи, костей и хрящей. Но ни один хищник не питается только мясом; в его рацион входит и растительная пища, причем
2. ОБОНЯТЕЛЬНАЯ ОРИЕНТАЦИЯ И ВОЗМОЖНОСТИ ОБОНЯТЕЛЬНОГО АППАРАТА
2. ОБОНЯТЕЛЬНАЯ ОРИЕНТАЦИЯ И ВОЗМОЖНОСТИ ОБОНЯТЕЛЬНОГО АППАРАТА Обонятельная система собак, связанная с первой парой черепно-мозговых нервов, морфологически легко отграничивается от других систем хеморецепции. Наиболее характерны различия между вкусом и обонянием.
Болезни опорно-двигательного аппарата
Болезни опорно-двигательного аппарата К болезням опорно-двигательного аппарата относятся заболевания костей, сухожилий, суставов, мышц. У лошадей часто регистрируются пороки конечностей, представленные в таблице 9, основным признаком которых является хромота.Таблица
Болезни опорно-двигательного аппарата
Болезни опорно-двигательного аппарата К болезням опорно-двигательного аппарата относятся заболевания костей, сухожилий, суставов и мышц. Все заболевания необходимо лечить под контролем ветеринарного
Болезни опорно-двигательного аппарата
Болезни опорно-двигательного аппарата К болезням опорно-двигательного аппарата относятся болезни костей, сухожилий, суставов, мышц. Все заболевания необходимо лечить под контролем ветеринарного врача.В данном разделе речь пойдет о некоторых распространенных
Болезни опорно-двигательного аппарата
Болезни опорно-двигательного аппарата К болезням опорно-двигательного аппарата относятся такие заболевания, как болезни костей, сухожилий, суставов, мышц. Любые заболевания опорно-двигательного аппарата необходимо лечить под контролем ветеринарного
Особенности дачного содержания молодой выставочной собаки
Особенности дачного содержания молодой выставочной собаки Уши, топ-ноты, всю гриву молодого выставочного пуделя или шерсть афгана, йорка или ши тцу необходимо защищать папильотками. Им категорически противопоказаны прогулки по высокой траве и кустарникам. Неизбежно
Устройства и сооружения
Устройства и сооружения Если вы разместили хомячка в клетке с подстилкой, положили ему еду, это ещё не значит, что он будет хорошо жить в такой клетке. Важно, чтобы клетка была оснащена разными устройствами, необходимыми хомячку для развития и полноценной жизни.Выше уже
БОЛЕЗНИ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
БОЛЕЗНИ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА Развитие скелета у собаки начинается еще в утробе матери и заканчивается в возрасте 9 – 12 месяцев и находится под контролем целого ряда гормонов, которые вырабатывают щитовидная и другие железы внутренней секреции. Гормоны