Генетика основных признаков у разных пород собак
Наследование пигментации
В собаководстве наиболее подробно разработана генетика окраски шерсти и пегости с многими вариациями, генетика типа шерстинок и шерстного покрова, генетика болезней аномалий.
Каждая порода характеризуется разной степенью изменчивости этих признаков и разными особенностями наследования.
В современной литературе (Робертсон, 1982) дано описание генетических особенностей 118 пород собак разного направления, для которых выявлено 29 аллелей пигментации. Так, например, по фенотипической и генетической изменчивости мастей наибольшая вариация выявлена у коккер-спаниеля (18 фенотипов), у пинчера (10 фенотипов), у пуделя (14 фенотипов).
Значительно меньшая изменчивость окраски шерсти наблюдается у собак сторожевого, служебного направления.
Несмотря на разнообразие окраски, можно выделить следующие основные ее типы и привести породы, имеющие в основном эту окраску:
Серая окраска — немецкая овчарка;
черная — ньюфаундленд;
кофейная (печеночного оттенка, коричневая) — доберман-пинчер;
голубая — доберман-пинчер;
соболиная, красная — боксер;
желтая, кремовая, светло-палевая — изабелла;
чепрачная — колли;
черная с подпалами — доберман-пинчер;
тигровая — дог, боксер;
альбинос (лейцисты); арлекины.
Такое разнообразие окраски происходит как в результате комбинативной и мутационной изменчивости, так и в результате наличия серий множественных аллелей.
Генетически обусловлено также распределение пигментации по телу. Различают собак одноцветных, двух- и трехцветных (сеттеры). Пигментация может быть от сплошной, пятнистой разной степени и типа рисунка до сильно выраженной утраты пигмента. Типы распределения пигмента сильно варьируют, а в ряде пород служат породным признаком.
Специфическая пятнистость выявлена у пойнтеров и сеттеров, у которых кроме обычной пятнистости в виде пегости имеет место так называемая крапчатость в форме мелких штрихо-образных или округлого типа черных, коричневых мелких пятен по белому фону. У гончих, кроме обычной чепрачности черного или коричневого пигмента, и у сеттеров описана «тиковая испещренность», когда белые волосы равномерно перемешаны с пигментированными. Она имеет доминантное наследование (ген F). Тиковая испещренность начинает формироваться у щенка к месячному возрасту.
Появление пятнистости затрагивает отдельные точки тела: голову, шею, хвост, лапы, крестец, спину. Депигментация реже обнаруживается на ушах и корне хвоста (лайки, фокстерьеры, гончие, овчарки). Считается, что появление белых пятен на ногах и спине имеет доминантное наследование, а на носу, ушах, бедрах — рецессивное.
Пигментация может распространяться по всей длине волоса (шерстинки) или в виде поперечно окрашенных разного Цвета зон (зонарная). Зонарная окраска распространена у немецких (восточноевропейских) овчарок, лаек и может сопровождаться чепрачным типом распределения по телу, в виде тигрового (полосатость) рисунка, с подпалами.
Чалость — (смесь неокрашенных волос с окрашенными) обусловлена доминантным геном (R). У догов с мышастой шерстью в генотип входит этот ген.
Вот некоторые данные, характеризующие фенотипические особенности пигментации собак разных пород:
Афганские породы — черные, черные с коричневым подпалом, голубые, серые;
борзые — черные, рыжие;
боксеры — рыжие, красные, тигровые;
бульдоги — красные, соболиные, тигровые;
чау-чау — черные, голубые, красные, серые;
доги — черные, коричневые, желтые, пестрые, арлекины;
доберман-пинчеры — черные, коричневые, голубые, изабелла;
японский хин — черные с белым, красные с белым;
ньюфаундленды — черные, печеночного цвета, голубые;
чи-хуа-хуа — 10 разных фенотипов.
Несмотря на большое фенотипическое разнообразие между породами и внутри пород, окраска собак, как показали генетические работы, обусловлена следующими основными генами и их аллелями.
Для того чтобы в организме мог синтезироваться пигмент, обусловливающий окраску шерсти, глаз, мочки носа, необходимо присутствие в генотипе гена С. Этот ген сам не вызывает окраску, а обеспечивает синтез пигмента как такового. При отсутствии доминантного гена С при его рецессивном аллеле с, даже при наличии аллелей, дающих черную, коричневую и другие окраски, собаки будут альбиносами, т. е. иметь белую шерсть, белую мочку носа, бесцветную радужную оболочку глаза (красные глаза), что наблюдается у полных альбиносов, но таких собак в практике собаководства не зарегистрировано. Но зато в ряде пород имеет место так называемый неполный альбинизм (лейцисты): при бесцветной (белой) шерсти сохраняется темная пигментация мочки носа и радужной глаз. Лейцизм зарегистрирован и считается породным признаком у белых шпицев, белых бультерьеров, у некоторых пород лаек.
Следовательно, все окрашенные собаки в генотипе имеют ген С и другие гены (черной, шоколадной, желтой пигментации).
Приведем перечень генов и их символы, обусловливающие разную окраску шерсти у собак.
Ген С — обеспечивает способность организма синтезировать пигмент любого цвета. При его отсутствии в переходе в рецессивный аллель С, наступает неполный альбинизм, несмотря на наличие генов, определяющих тот или иной цвет.
Ген А — определяет зонарное распределение пигмента вдоль шерстинки и дает окраску типа «агути», характерную для диких животных этого семейства, его рецессивный аллель а приводит к отсутствию зонарности.
Ген В — в доминантном состоянии обеспечивает синтез черного пигмента, а его рецессивный аллель b дает коричневый (кофейный) цвет.
Ген Е — определяет полное распространение черного или коричневого пигмента по шкуре, а его рецессивный аллель е обеспечивает синтез желтого и красного пигментов.
Ген S — дает доминантную сплошную окраску шерстного покрова, рецессивный аллель s — пятнистость.
Ген D — усиливает интенсивность пигмента в корковом и мякотном веществе волоса, рецессивный аллель d приводит к ослаблению пигментации, переводит черный цвет в голубой.
Ген ер — обусловливает тигристость.
Ген W — определяет доминантную белую окраску шерсти.
Ген Н — определяет доминантную окраску типа «арлекин».
Ген cd— ослабитель красного цвета до желтого.
Ген R — чалая окраска шерстного покрова.
Ген Т — тиковая пятнистость.
Учитывая действие указанных основных генов пигментации, приведем в качестве иллюстрации полные генотипы собак определенной окраски и породной принадлежности (по Н. А. Ильину).
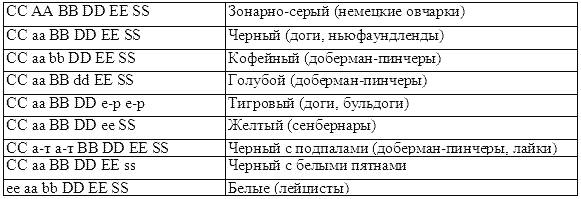
В последнем типе, у лейцистов, гены окраски находятся в скрытом (криптомерном) состоянии и не могут проявить свое действие, поэтому собаки-лейцисты — белые. Но при скрещивании белой собаки-лейциста с окрашенной собакой, у которой есть ген С, в потомстве могут быть как окрашенные, так и лейцистные щенки, т. е. с черной, кофейной и альбиноской окраской.
С окраской шерсти коррелирует часто и пигментация глаз. Различают карие, коричневые, желтые, голубые, голубовато-белесые, резко рубиновые (из-за отсвечивания зрачка). Иногда наблюдается разноглазость по цвету радужной оболочки у одной и той же особи, что встречается у арлекинов и выявлено у догов и гончих.
Ген У. Обусловливает желто-коричневую радужную глаза, его рецессивный аллель У, дает голубые глаза.
Ген Р. Доминантное состояние нормального глаза. Рецессивный ген Ра, дает рубиновые глаза, что выявляется при определенном положении глаза и повороте головы. Эта окраска может сочетаться со светлой окраской радужной (белый глаз). Такие глаза встречаются у собак разных пород. Н. А. Ильин зарегистрировал эту особенность у 12 пород: доги, сеттеры, лайки, курцхаары, боксеры, гончие, эрдельтерьеры, немецкие овчарки, белые шпицы, таксы и др. Рубиновоглазие может сочетаться с нормальной окраской другого глаза этой же собаки. Такое явление в пигментации глаз зарегистрировано у человека, кошки, кролика, мыши, крысы. Не следует смешивать это с красноглазием при альбинизме, так как при нем пигмент отсутствует и в радужной глаза.
Наследование некоторых элементов экстерьера
Генетически изучаются и другие элементы экстерьера: тип и структура шерстного покрова, форма и длина ушей и хвостов, особенности строения костей черепа.
Различают следующие типы шерстного покрова и гены, их обусловливающие.
Нормальная короткая шерсть (ген l — доберман-пинчеры, боксеры и др.).
Длинношерстные (ген i — немецкие овчарки, ньюфаундленды, колли).
Иглокороткошерстные, жесткошерстные (ген r — фокстерьеры, жесткошерстные легавые).
Шелковистые (болонки), бесшерстные (ген Н — голые мексиканские).
Прямой волос.
Полуволнистый волос (пудели).
Завитковый волос (пудели).
Скрещивание короткошерстной собаки, несущей в генотипе доминантный ген L, с длинношерстной собакой, имеющей рецессивный ген I, часто дает в потомстве не полное, а промежуточное наследование длины шерсти: в приплоде будут потомки с типом волоса, отклоняющегося в той или иной степени в сторону одного или другого родителя. Это объясняется тем, что длина волоса является количественным признаком и обусловлена влиянием многих генов, то есть имеет полигенное наследование.
Наследование формы и размера ушной раковины также обусловлено генетически несколькими аллелями. Установлено, что полустоячее ухо (ген На) характерно, например, для собак породы колли, фокстерьеров, которые могут иметь генотипы На, НаН, Hah.
Висячее ухо (ген R) с генотипом НН распространен у многих пород (спаниели, гончие, таксы), стоячие — ген h с генотипом hh (немецкая овчарка). Часто наблюдаются промежуточные формы и размеры ушей. Наследование ушной раковины подчиняется влиянию нескольких однозначных генов, то есть имеет тип полимерного наследования. Условия выращивания и климатические факторы тоже могут оказывать влияние на этот признак, так как эти факторы влияют в определенной мере на формирование конституции, появление рыхлости, или наоборот, излишней сухости и переразвитости.
Характерным наследственным и породным признаком служит длина и форма хвоста. Различают породы длиннохвостые, со средней длиной хвоста (до скакательного сустава), короткохвостые и бесхвостые (полное отсутствие хвостовых позвонков). Наследование длины хвоста обусловлено полимерией. Поэтому скрещивание длиннохвостой с короткохвостой собакой дает в их потомстве собак с варьированием длины хвоста. Однако в практике собаководства наблюдаются случаи рождения короткохвостых собак с уменьшенным числом позвонков.
Считают, что на фенотипическую длину хвоста оказывают влияние гены-модификаторы и в некоторой степени внешние факторы.
Прирожденная короткохвостость, появляющаяся изредка у отдельных особей, была использована в селекции и привела к созданию новых пород: шиперке (карликовый шпиц корабельщиков), гладкошерстная легавая бурбон и др.
Форма хвоста и его постав наследственны и закреплены человеком путем селекции. При экстерьерной оценке собак на рингах этим особенностям придается важное значение как элементу, характеризующему породу. Например, эти требования распространяются на лаек (хвост кольцом на спине), фокстерьеров и эрдельтерьеров (прямостоячий хвост), легавых (горизонтальное положение хвоста прутом) и др.
Большое разнообразие у собак имеет строение черепа, часто вызванное мутацией и закрепленное селекцией, как породный признак. В результате доместикации(одомашнивания) и искусственного отбора и подбора основные морфологические особенности костей черепа, типичные для волка и дикой собаки, претерпели изменения. Удлиненные кости черепа, особенно челюстного аппарата, типичны для собак борзых пород, мощный и тяжеловесный череп характерен для ряда сторожевых собак, мопсовидность с крайним проявлением в строении черепа наблюдается у различных пород болонок, мопсов и т. п. Мутационные изменения в строении скелета конечностей (например, коротконогость), являющиеся для дикого вида собачьих пороком, были закреплены человеком в качестве породного признака у такс, что позволяет собаке удобнее работать в норных условиях охоты.
Прибылые пальцы, в виде пятого пальца на задней ноге, собак разных пород также наследственно обусловленный признак. В практике собаководства их обычно удаляют у щенков при рождении, чтобы предотвратить возможные травмы у взрослого животного.
Таким образом, для каждой породы собак экстерьер и конституция типичны и обусловлены наследственностью, которая основывается на полигенном, плейотропном действии гена.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК